|
Home >Recent Advances .
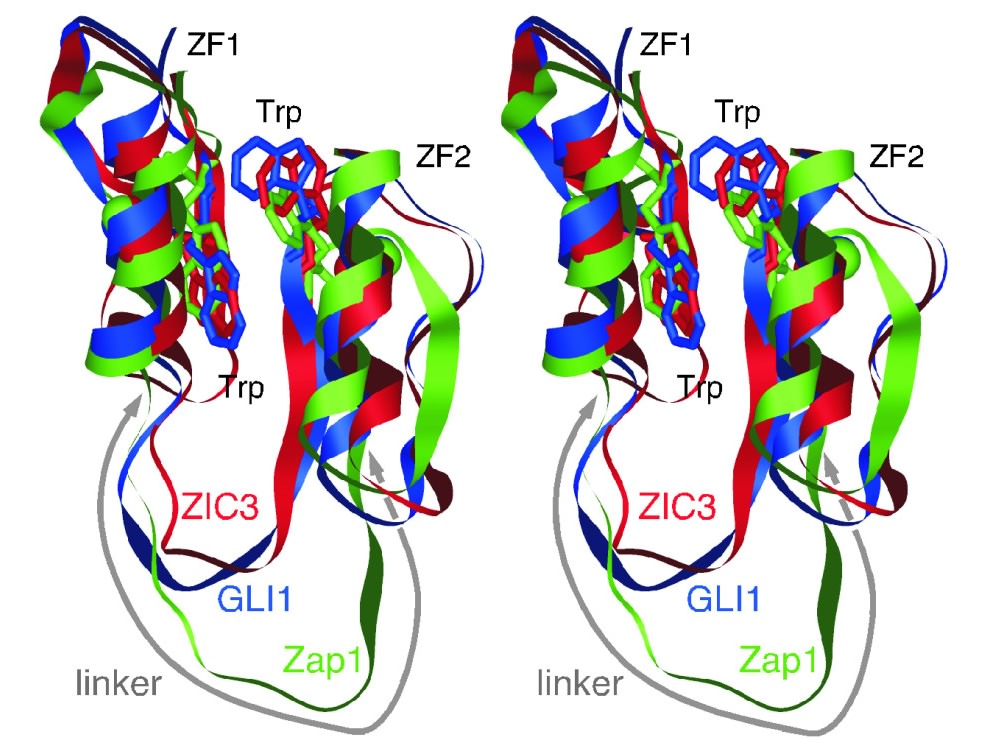
Discovery of a hallmark sequence motif for inter-zinc finger interaction (Hatayama et al., 2010) The C2H2 zinc finger (ZF) domain is widely conserved among eukaryotic proteins. In Zic/Gli/Zap1 C2H2 ZF proteins, the two N-terminal ZFs form a single structural unit by sharing a hydrophobic core (Fig. 1).
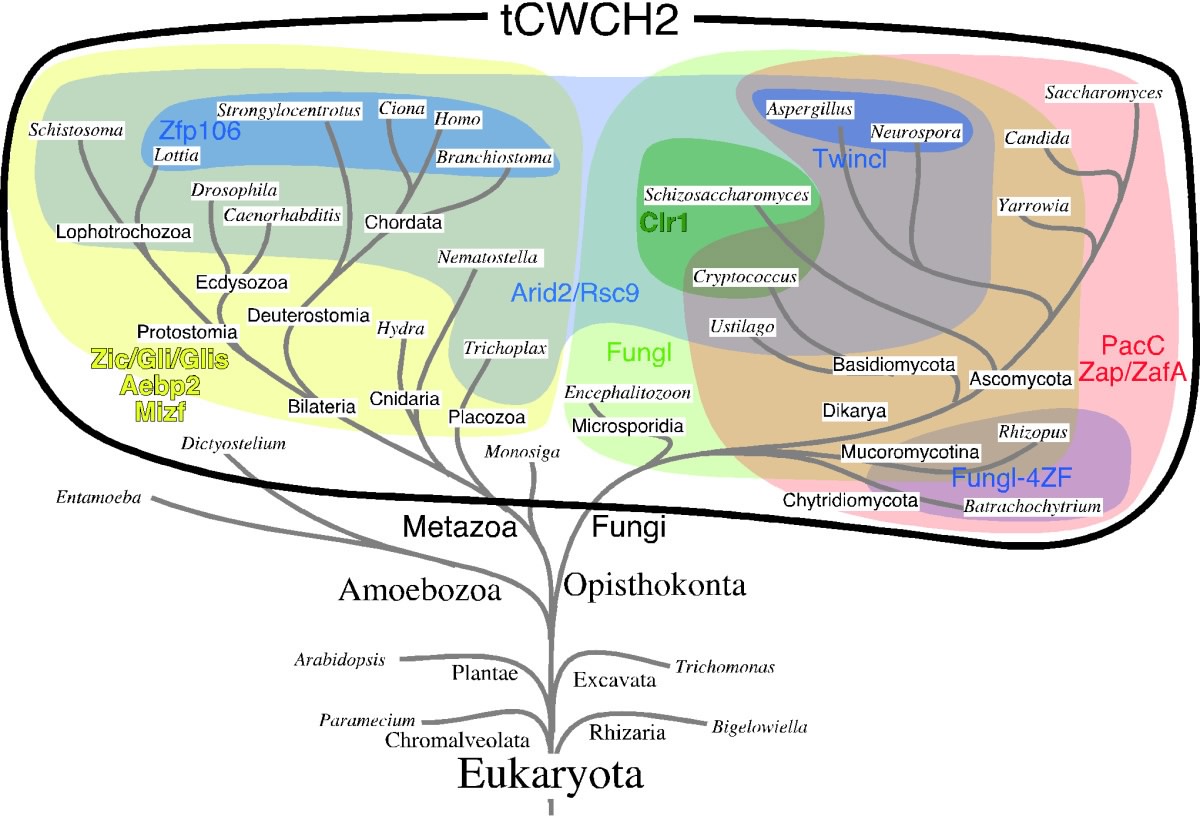
Fig. 1 Structures of tCWCH2 sequence motifs from Zic/Gli/Zap1 This structural unit defines a new motif comprised of two tryptophan side chains at the center of the hydrophobic core. Because each tryptophan residue is located between the two cysteine residues of the C2H2 motif, we have named this structure the tandem CWCH2 (tCWCH2) motif (See also "Clarification of the 3D structure of the interspersed nuclear localization signal in zinc finger domain"). In this study, we characterized 587 tCWCH2-containing genes using data derived from public databases. We categorized genes into 11 classes including Zic/Gli/Glis, Arid2/Rsc9, PacC, Mizf, Aebp2, Zap1/ZafA, Fungl, Zfp106, Twincl, Clr1, and Fungl-4ZF, based on sequence similarity, domain organization, and functional similarities. tCWCH2 motifs are mostly found in organisms belonging to the Opisthokonta (metazoa, fungi, and choanoflagellates) and Amoebozoa (amoeba, Dictyostelium discoideum) (Fig. 2). By comparison, the C2H2 ZF motif is distributed widely among the eukaryotes.
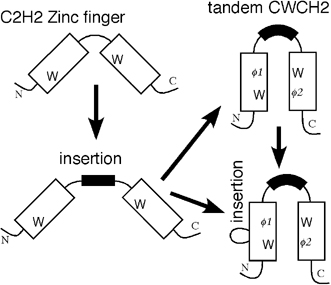
Fig. 2 Distribution of tCWCH2-containing genes in a eukaryotic phylogenetic tree The structure and organization of the tCWCH2 motif, its phylogenetic distribution, and molecular phylogenetic analysis suggest that prototypical tCWCH2 genes existed in the Opisthokonta ancestor. Within-group or between-group comparisons of the tCWCH2 amino acid sequence identified three additional sequence features (site-specific amino acid frequencies, longer linker sequence between two C2H2 ZFs, and frequent extra-sequences within C2H2 ZF motifs). These features may be explained by adaptation of its role in mediating the inter-ZF interaction (Fig. 3).
Fig. 3 Evolution of the tCWCH2 domain. We propose that the tCWCH2 patterns were generated from classical C2H2 sequences concurrently with the acquisition of the additional sequence features These results suggest that the tCWCH2 motif is a specialized motif involved in inter-zinc finger interactions.
|