|
トップページ >主な研究成果
Zicタンパク質によるWntシグナルの抑制が初期発生に重要であることを発見 (Fujimi et al., 2012)
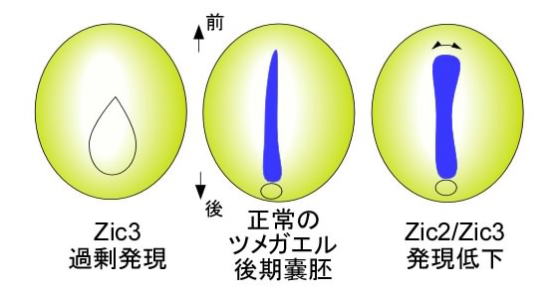
アフリカツメガエルは動物の初期発生の研究材料として非常に優れています。私たちはアフリカツメガエル胚を用いた実験を行い、初期発生におけるZicファミリータンパク質の重要な機能を明らかにしてきました。これまでにZicファミリータンパク質が、神経板・神経堤が外胚葉から分化する過程で重要な役割を持つこと、中胚葉の発生においても重要な役割を持ち、それが体の左右非対称性と関わること等が分かっています。 アフリカツメガエルでは受精に引き続いて、卵割、胞胚、嚢胚、神経胚、尾芽胚と各発生時期に特有な外見を示し、形態が大きく変化していきます。このうち、胞胚後期から嚢胚初期にかけて原腸陥入(嚢胚形成)という過程を経ますが、この過程で、前後、左右、背腹に沿ってどのような組織がどのように形づくられるかといった大まかなプランが決定されます。このうち、脊索は背中側にある棒状の構造で、発生過程にだけ認められます。これは、哺乳類、鳥類、は虫類、両生類、魚類を含めた脊椎動物とホヤやナメクジウオなど脊索動物に特徴的な構造です。 この研究の中で私たちは、Zicタンパク質が脊索の発生に対して抑制的な働きを持つことを見いだしました。Zic3を過剰に産生させると、脊索のマーカー遺伝子(Xnot)の発現が部分的に失われます(図1左)。一方、Zic2とZic3の量を減少させると、マーカーの分布が左右に広くなります(図1右)。もう少し発生の進んだ段階の胚では、Zic3を過剰に産生させた胚では背側の脊索が失われており、Zic2とZic3の量を減少させた胚では前方の脊索が太くなることが分かりました。
図1 背中側でZicの発現(産生)量を変えた胚での脊索マーカーの分布(青) 発現状態の変化は卵割期の胚にメッセンジャーRNAまたはその働きを抑えるRNA(アンチセンスRNA)を注入することにより引き起こしました。 これまでに脊索が形成される過程で、Wnt-βカテニン経路が重要な役割を持つことが知られていました。細胞外に分泌されるWnt(ウイント)タンパク質がWnt受容体に働きかけ、それがいくつかの分子の変化を引き起こし、βカテニンという核内で遺伝子のスイッチを切り換えるタンパク質の状態を変化させます。Zicタンパク質はこのWnt-βカテニン経路の情報を抑えていることが明らかになりました(図2)。
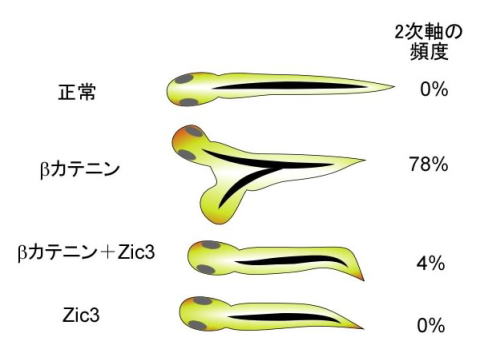
図2 Zicがβカテニンの働きを抑制することを示す実験の一つ オタマジャクシを背側からみたところ。βカテニンをアフリカツメガエルの胚の腹側で異所的に発現させると本来の脊索以外にもう一つよけいな脊索(2次軸)が形成されます。このときにβカテニンと同時にZic3を発現させると、2次軸の形成が抑えられます。 Wnt-βカテニン経路は動物の発生過程や腫瘍形成のさまざまなところで働いています。おそらくZicとWnt-βカテニン経路の関係は脊索の調節だけにとどまらないだろうと私たちは予測しています。今後ZicによるWnt-βカテニン経路の抑制活性をどのように応用することができるかについても検討をしていきたいと考えています。
|