|
トップページ >主な研究成果
抑制性シナプス形成に重要なタンパク質を発見 (Takahashi et al., 2012)
○膜タンパク質SLITRK3が抑制性シナプスの形成を誘導 ○SLITRK3欠損マウスはてんかんに似たけいれん発作を起こす ○神経細胞の過活動と関連した神経疾患の病態の理解や改善に期待
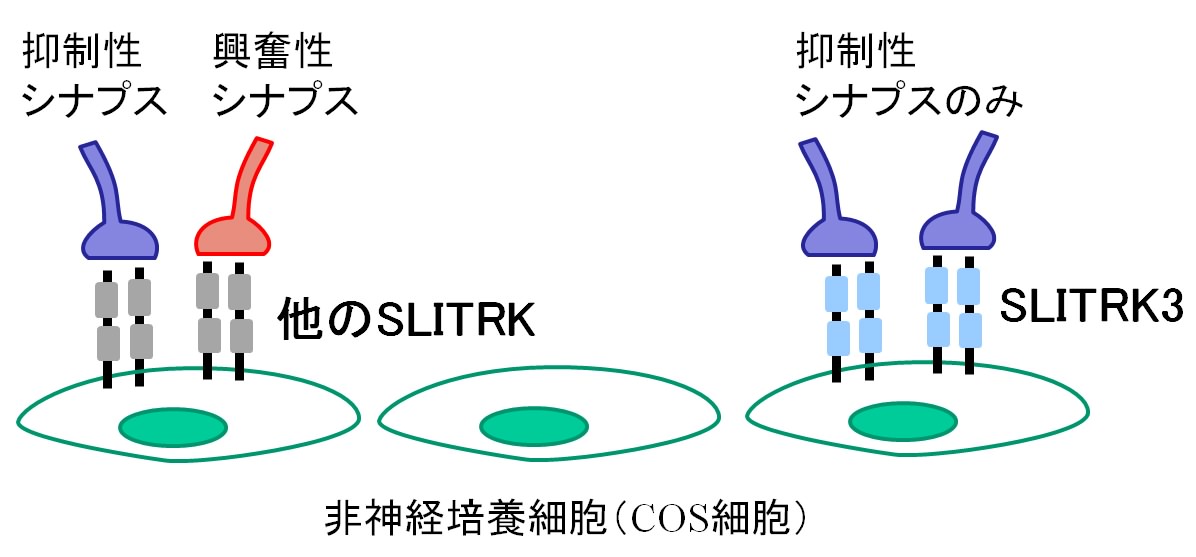
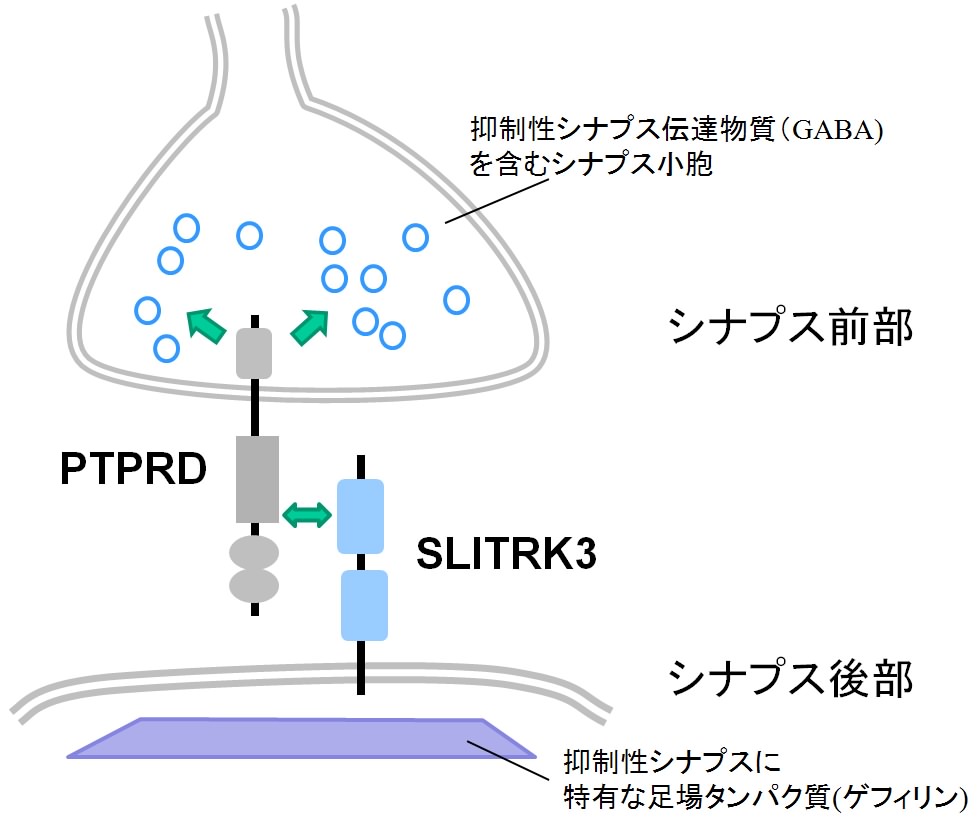
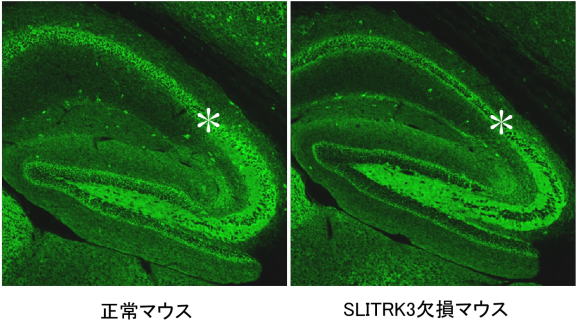
神経細胞は情報を伝達することに特化した特別な細胞で、動物の感覚や行動などを制御しています。神経細胞同士が情報を伝え合うときには、神経細胞間に形成されるシナプスという構造が重要な役割を果たします。情報の伝達に神経伝達物質が使われるタイプのシナプスは、機能的に「興奮性シナプス」と「抑制性シナプス」に分けることができます。興奮性シナプスでは情報を受け取った細胞が電気的に興奮やすくなり、抑制性シナプスでは興奮しにくくなります。近年の研究から興奮性シナプスと抑制性シナプスのバランスが正常な脳機能活動に重要で、いくつかの神経疾患ではこのバランスが乱れていると考えられています。 最近の研究により、シナプスの形成に関わる分子について理解が進み、興奮性シナプスの形成メカニズムについては多くのことが分かってきました。しかし、抑制性シナプスの形成については、これに関わる分子が少数しか見つかっていないため、あまり理解が進んでいませんでした。 SLITRKファミリーは、私たちが発見、命名した神経系に発現する細胞膜貫通型タンパク質ファミリーです。6種類の細胞膜貫通型タンパク質からなるSLITRK1~SLITRK6が脳神経系や内耳などで産生されること、一部のものは高次脳機能の調節、神経回路の形成に重要な役割を持つことを報告しました(内耳の神経回路形成とSLITRK6)。 2009年と2011年に米国とカナダのグループがこのタンパク質ファミリーとヒトとの精神神経疾患への関与の可能性を報告し、疾患原因遺伝子の候補として注目を集めています。そこで私たちは、SLITRKファミリーとシナプスの関わりに着目し、その機能や分子機構の解明に挑みました。 始めにSLITRKファミリーとシナプス形成の関係性を調べるために6種類のSLITRKファミリータンパク質をそれぞれ発現させた非神経細胞とラットの海馬の神経細胞を混ぜ合わせて培養しました。すると、SLITRKファミリータンパク質を発現させた細胞に対して、海馬の神経細胞がシナプスに似た構造を作ることが明らかになりました。興奮性シナプスと抑制性シナプスそれぞれの局在が分かる特有な分子マーカーで調べると、SLITRK3タンパク質以外を発現させた非神経細胞は、興奮性と抑制性シナプス両方を同程度に誘導するのに対して、SLITRK3タンパク質を発現させた非神経細胞は、抑制性シナプスだけを強く誘導することが分かりました。 (図1)。 図1 培養細胞で見いだされたSLITRK3のシナプス形成活性 また、培養した海馬の神経細胞では、SLITRK3タンパク質は抑制性シナプスのシナプス後部(神経伝達物質を受け取る側)に存在する足場タンパク質(ゲフィリン)と共在し、興奮性シナプスの後部に存在する足場タンパク質(PSD95)とは共在しないという結果を得ました。さらに海馬の培養細胞の中で、SLITRK3タンパク質の遺伝子発現を減少させると、ゲフィリン陽性の抑制性シナプスの数が減少するのに対して、PSD95陽性の興奮性シナプスの数には、明確な影響がありませんでした。つまり、SLITRK3タンパク質は抑制性シナプスを誘導し、シナプス後部にゲフィリンと共在していることが分かりました。 シナプス形成には、シナプスの前部と後部の間でいくつかの分子が結合され、分子複合体を形成することが必要です。そこで、これまでにシナプスの形成に関与することが知られているいくつかの分子について、SLITRK3と結合するかどうかを検討しました。その結果、膜タンパク質の一つであるPTPRDがSLITRK3と結合することを見いだしました。PTPRDはシナプス前部(神経伝達物質を放出する側)に分布しており、培養した海馬の神経細胞でPTPRDの遺伝子発現を減少させると、SLITRK3を発現させた非神経細胞と混合培養しても、抑制性シナプスが形成されにくくなりました。つまり、SLITRK3はPTPRDと結合して、抑制性シナプス前部の構造を誘導すると考えられます(図2)。 図2 SLITRK3とPTPRDの結合が抑制性シナプスの形成を促進する SLITRK3はシナプス後部に、PTPRDはシナプス前部に局在している。両者の結合が抑制性シナプスを構成する分子複合体の形成を促進するものと考えられる。 実際に脳の中でSLITRK3が抑制性シナプス形成に関与するのか調べるために、SLITRK3欠損マウスを作製し、そのシナプスの異常を調べました。すると、海馬の錐体細胞の細胞体周囲で抑制性シナプス特有の分子マーカーの発現が減少していました(図3)。そこでSLITRK3欠損マウスの海馬の切片でシナプスの活動を調べると、抑制性シナプスの活動に由来する電気信号が減少するのに対して、興奮性シナプスの活動に由来する信号には変化が認められませんでした。つまり、SLITRK3欠損マウスの海馬の錐体細胞では、抑制性シナプスが減少することを確認しました。 図3 SLITRK3欠損マウスの海馬での抑制性シナプスの様子 SLITRK3欠損マウスの海馬の錐体細胞層(*印)で、抑制性シナプスに特有な分子マーカーに対する抗体を用いたところ、正常マウスに比べて蛍光色が少なくなっている。つまり、抑制性シナプスが顕著に減少していることを示している。 SLITRK3欠損マウスは、時々てんかんの発作に似た異常行動(突然無動状態になる、頭部を規則的にゆする、前肢を硬直させるなど)を示します。また、脳波を調べると振幅の大きい異常波が頻繁に観察できました。これらの異常は、おそらく抑制性シナプスの機能が失われたために起きる神経細胞の過活動が原因であると推測できます。 今回、神経系に発現する膜タンパク質SLITRKファミリーのひとつのSLITRK3が、神経細胞の抑制性シナプスの形成に重要な役割を持つことを発見しました。今後、抑制性シナプスの脳機能における役割を詳細に解明するためにSLITRK3欠損マウスの神経回路の異常や行動の異常を検証します。特にSLITRK3を含めたSLITRKファミリーがもつ中枢神経系の興奮性・抑制性シナプスのバランスを制御する仕組みの解明は、基礎研究の観点から大変重要なテーマです。さらにSLITRK3を含む抑制性シナプスの形成・維持に関わる分子機構を解明できれば、てんかんや多動症など神経細胞の過活動と関連した神経疾患の病態の理解や改善に役立つと期待できます。 <補足説明> 神経系に発現する新規の細胞膜貫通型タンパク質ファミリーとして、2003年理研脳科学総合研究センターの発生発達研究グループにより報告された。1回膜貫通型タンパク質で、アミノ末端側にはロイシンリッチリピートというタンパク質-タンパク質間の結合によく用いられるドメインが2個存在し、カルボキシ末端側には神経栄養因子受容体と部分的に類似のアミノ酸配列が存在する。SLITRK 3は脳全体で発現するが、特に海馬の錐体細胞層などで強いことが知られている。
神経細胞間あるいは筋繊維、または神経細胞と他種細胞間に形成される、シグナル伝達などの神経活動に関わる接合部位とその構造のこと。化学シナプス(小胞シナプス)と電気シナプス(無小胞シナプス)、および両者が混在する混合シナプスに分類される。シグナルを伝える側の細胞をシナプス前部、伝えられる側の細胞をシナプス後部と呼ぶ。神経細胞の増殖の際に、特定の位置にある神経細胞が特定の細胞に軸索を伸ばし、シナプスを形成して神経回路を形成する。そのときに軸索を誘導する因子として神経栄養因子(ニューロトロフィン、NGFなど)が知られている。また、神経細胞群は初期に過剰な接続を形成した後、必要なものだけを残してシナプスを減らすと考えられている。
受容体型チロシンホスファターゼ。膜貫通型のタンパク質であり、通常2個のホスファターゼ関連ドメインを有している。細胞外はさまざまな分子間相互作用に関わる免疫グロブリン様ドメイン、フィブロネクチン3様ドメインが存在している。このグループのホスファターゼは、細胞外の情報を細胞内の伝達経路に伝える役割を担っていることが多い。PTPRD欠損マウスは空間学習機能に異常があり、海馬のシナプスの性質が変化していることが知られている。
神経伝達物質に対する受容体や様々な情報伝達物質が分子複合体を形成する足場となるタンパク質。ゲフィリン(GEPHYRIN)は、抑制性シナプスのシナプス後部に特有に存在するタンパク質で、PSD95は興奮性シナプスのシナプス後部に特有に存在する足場タンパク質であることが知られている。 海馬に存在する主要な興奮性の神経細胞。その細胞体は明瞭な層状構造を形成する。数多くの海馬の抑制性神経細胞が錐体細胞の様々な部位に抑制性シナプスを形成して、その活動を制御していることが知られている。 この研究は カナダブリッティシュコロンビア大学の高橋秀人、Tuhina Prasad、Ann Marie Craig、理化学研究所脳科学総合研究センター吉原良浩チームの宮本浩行、同センター津本忠治チームの惣谷和広、津本忠治、群馬大学の安田浩樹との共同研究です(敬称略)。
|