
試験管内遺伝子転写機構
数十キロベースの鋳型を用いてクロマチン形成と試験管内遺伝子転写を行っています.この試みは長崎大学医学部生化学教室が世界的にもリードしている分野です.基礎配属の学部学生に試験管内転写の手技を教授しながら、助教中川武弥がよりin vivoに近い遺伝子転写系の確立を行っています.
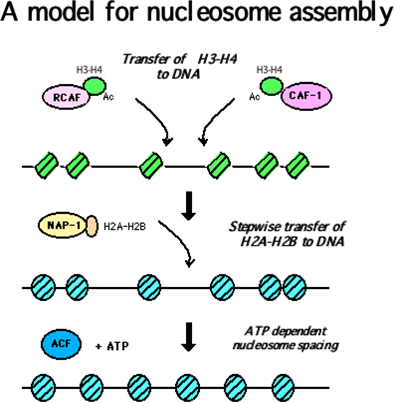
生化学教室ではクロマチン形成因子NAP-1とACFによるクロマチン形成の系を用いて、この順番がヒストン結合タンパクであるNAP-1、およびDNAとH2A-H2Bの結合力がヒストンH3-H4のDNAへの転位により変化することをみいだしました.さらにヌクレオソーム形成に引き続き、ヌクレオソームのスライドが起きることを明らかにしました(JBC276,2001).すなわち我々は下図Nucleosome assemblyモデルを証明し、またこの方法は共同研究者に教授しています.またこのヌクレオソームはACF の存在下に、遺伝子転写の際に起きる再構築能を有しており、クロマチン構成因子ACF はクロマチン形成と再構築の両現象に関与していることをみいだしました(Cell 90, 1997).
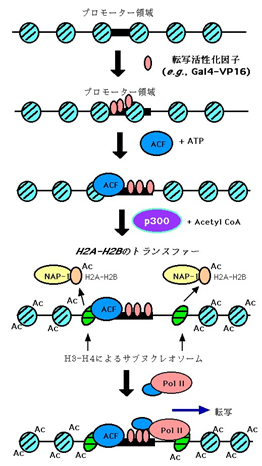
ヌクレオソームの再構築は遺伝子転写の初期の現象としてとらえられています.この現象に関与する因子として、ATP依存性のクロマチン再構築因子に加え、近年ヒストンのアセチル化酵素が注目されている。転写活性化因子のコアクチベーターとして知られるp300等がヒストンアセチルトランスフェラーゼ(HAT)活性を持ち、ヒストンをアセチル化することによって、特異的な遺伝子転写の調節に関与することが知られるようになったからです.しかし、アセチル化により何が引き起こされるのか実際に証明されていませんでした.我々の系を使ってp300によるアセチル化は転写活性化因子に依存して起きることを明らかにしました.さらに転写活性化因子(GAL4-VP16)はp300によるヌクレオソームコアヒストンのアセチル化を促進するがクロマチンのリモデリングはヒストンのアセチル化とは独立して起こることを明らかにしました.またACFとATPはp300によるコアヒストンのアセチル化に重要であることを明らかにした。この結果から、ATPとACFを必要とするクロマチンリモデリングは転写反応の最初のステップであり、引きつづいて起きるコアヒストンのアセチル化を制御していると考えられます.(Genes Dev 14, 2000)。すなわち我々は下図遺伝子転写開始を証明しました.
(文責;長崎大学医学部生化学教室 伊藤敬)