
運動の制御を行う線条体で酵素や受容体の分布を決めるタンパク質を発見
Ichise et al., Front Mol Neurosci. (2022)
Lrtm2は線条体投射システムにおけるタンパク質の選択的な輸送を制御し、その欠乏は運動機能とモノアミン動態の異常を引き起こす
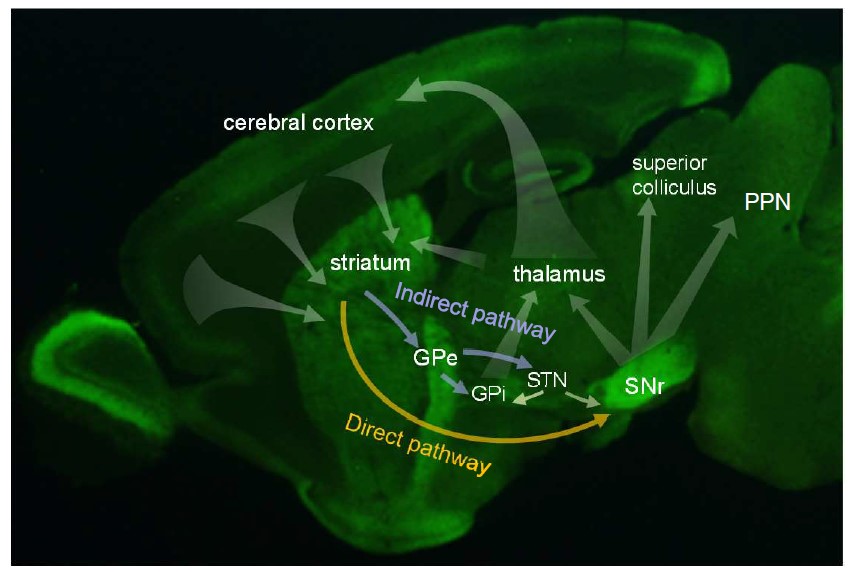
脳の線条体(striatum)は大脳の深いところに位置している大脳基底核の一部で、運動の制御に関与しており、運動の制御にかかわることが、知られています。線条体は体性感覚野や前頭前皮質などの大脳皮質(cerebral cortex) などからの入力を処理して、黒質網様部(SNr)や淡蒼球(GPe)へ出力します。出力を担うのは、線条体の中にある中型有棘神経細胞で、黒質網様部、淡蒼球に軸索(アクソン)を伸ばしています。(下図)。
この出力に関わる中型有棘神経細胞の軸索末端部(神経終末)には神経伝達物質GABAを合成するGABA合成酵素(GAD67)とフィードバックを受けるGABA受容体(GABABR1)が存在しています。これらは適量のGABAが神経終末から放出ために重要で、正常に働くためには神経細胞の中で正しい場所へ運ばれる必要があります。私たちはこの研究でLrtm2という膜貫通タンパク質がこの輸送に関わっていることを明らかにしました。
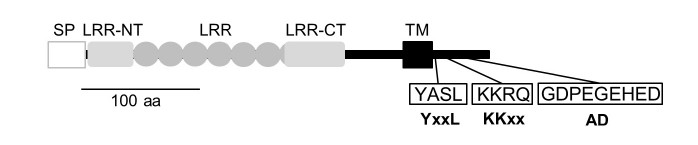
Lrtm2(下図)はロイシンリッチリピートドメインを持つ脳の膜貫通タンパク質の一つで、マウス全身の臓器の中では脳に強く発現し、脳内では線条体の中型有棘神経細胞とその投射先である黒質網様部(SNr)、淡蒼球、線条体に強く分布しています(上の図の緑色の部分)。また、細胞の中では、軸索突起、シナプス、シナプス小胞などにあります。
Lrtm2遺伝子を欠損したマウスの線条体では、GAD67とGABABR1が正しく輸送されなくなっていました。そこで、Lrtm2には線条体の中型有棘神経細胞の中で酵素や受容体の細胞内局在を決める性質があるのではないかと考え、軸索突起に運ばれない変異型Lrtm2タンパク質を作製しました。この変異型Lrtm2タンパク質は培養した神経細胞において、GAD67およびGABABR1を軸索に運ぶ能力が正常のLrtm2よりも低下していました。また、Lrtm2 欠損マウスの脳においても変異型Lrtm2は正常Lrtm2よりもGABABR1を運ぶ能力が低下していました。
先に説明したように、線条体は、運動の制御に関わる領域です。そこで、Lrtm2欠損マウスに運動機能の異常がないか確かめてみました。すると、Lrtm2欠損マウスは、新たなマウスや物体に出会ったとき、回転する棒の上に留まるとき、水槽に入れられたときなど、様々な環境に置かれたときの行動選択に異常を示しました。また、線条体の機能維持に重要な役割を果たす線条体のドパミンの代謝産物量が変化していることがわかりました。これらの結果はLrtm2の欠損が線条体の機能に影響を及ぼしたことを示唆しています。
以上の結果から、脳機能におけるLrtm2の役割は、随意運動の開始、運動学習の調節、探索行動の制御であり、この役割には線条体-黒質投射系に局在しているLrtm2が関わっているものと考えられました。
今後、Lrtm2を含む分子装置の実体を明らかにすることにより、神経細胞の軸索輸送の機構および軸索終末部の機能調節の機構についてより良い理解ができ、線条体の機能異常を伴う神経疾患の病態の理解にも貢献できるものと期待しています。
