
初期胚の細胞運命決定に関わるゲノム要素
「Zic3中胚葉エンハンサー」の発見
Odaka et al., Sci Rep.8, 14954 (2018)
ZIC3は内臓錯位症候群 (左右不定位症)およびそれに関連した心奇形やVACTERL連合と呼ばれる奇形症候群の原因遺伝子の一つです。Zic3遺伝子から作り出されるZic3タンパク質は、セキツイ動物の幹細胞で細胞の運命決定に重要な役割を持つことが明らかになっています。
受精卵は細胞分裂を繰り返して、やがて外胚葉・中胚葉・内胚葉と3つの領域に分かれます。中胚葉は将来、筋肉や骨などを作り出しますが、中胚葉由来の細胞の運命決定の過程でZic3タンパク質が働いていることが以前の研究で明らかになっています(Zicタンパク質によるWntシグナルの抑制が初期発生に重要であることを発見, Fujimiら, 2012)。しかし、どのようにして中胚葉でZic3遺伝子の情報が読み出されるようになるのか(Zic3遺伝子が発現するのか)についてはよく分かっていません。
この研究ではZic3遺伝子の中胚葉での発現に重要な役割をもつゲノムDNA中の領域(エンハンサー)をニワトリやマウスの胚を用いて探索しました。そして、見いだされたエンハンサーにどのような分子が結合して、細胞の運命を決定しているのかを調べました。
まず、Zic3遺伝子周辺のDNA塩基配列をニワトリとマウスで比較し、タンパク質をコードしていない領域で、特に良く保存された(配列が似ている)領域を選び出しました。このような領域にはエンハンサーが存在する可能性が高いことが、これまでの研究で知られていたためです。次に、これらの領域を、蛍光タンパク質(GFP)をコードするDNAの上流に配置した人工遺伝子(レポーター遺伝子)を作成しました。このレポーター遺伝子を、ニワトリの初期胚に培養下で導入してGFPの発現の度合いを調べ、エンハンサーの働きをもつ領域をいくつか特定しました(下図)
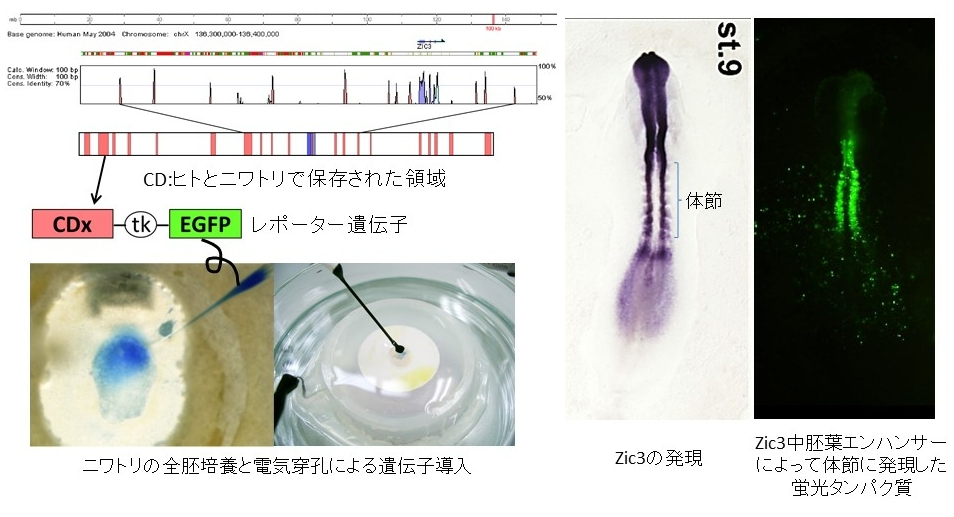
上の実験で見いだされたエンハンサー候補について、次にマウスで同じような実験をおこないました。保存された配列に相当する部分をマウスのゲノムDNAから取り出し、新たにレポーター遺伝子を作成し、マウスの胚に導入したところ、いずれの保存配列もニワトリの胚と同じ場所に発現を導く傾向がありました。その中で特にはっきりと初期胚の中胚葉組織における発現を導く保存配列を選び、この配列を欠失するマウスを遺伝子相同組み換えによって作成しました(中胚葉エンハンサー欠損マウス)。このマウスにおけるZic3の発現を調べたところ、初期中胚葉やそこから生じる体節と呼ばれる構造にZic3の発現がなくなっていることがわかりました(下図)。これらの結果から私たちの同定した中胚葉エンハンサーはニワトリとマウス(を含むおそらく広い範囲の脊椎動物)の間で、共通のZic3発現調節領域であることがわかります。

Zic3の体節における発現が消えている。
それではこの「中胚葉エンハンサー」はどのようにしてZic3の発現を制御しているのでしょうか。遺伝子の発現は、特定のDNA配列を認識して結合する転写因子に分類されるタンパク質が大きく関わっています。私たちはクロマチン免疫沈降(ChIP-seq)という実験のデータベースを利用して、どのような転写因子が「中胚葉エンハンサー」に結合するのか調べてみました。それによると中胚葉への分化に重要な役割を持つことが知られているBrachyury (T)やWntシグナルの下流因子が結合する他に、ES細胞の多分化能維持に重要であることが知られているOc3/4やNanog、初期胚の細胞運命決定に重要な役割を持つZic2が結合することが判明しました。(下図)。
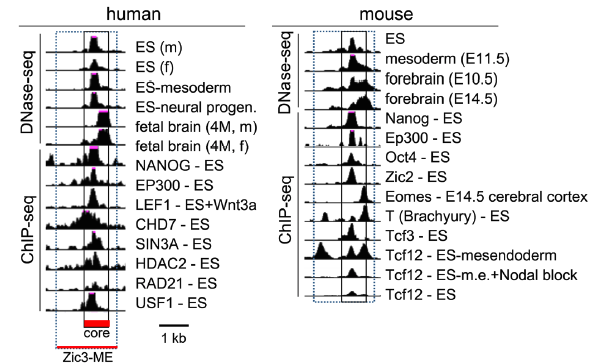
Zic3中胚葉エンハンサー(Zic3-ME)の中で
どこがどの時期に活性化しているか(DNA-seq)、
どこにどのタンパク質が結合しているか(ChIP-seq)を示す。
ここから、Zic3の「中胚葉エンハンサー」は初期胚の細胞運命決定でスイッチの役割を果たすことが予想されました。ES細胞に内胚葉へと運命づける「Nodalシグナル」が活性化するとZic3の中胚葉エンハンサーにTcf12という転写因子が結合するようになり、Tcf12はZic3の発現を抑制します。これまでの研究から、Zic3タンパク質はES細胞が内胚葉になることを抑える性質を持つことが知られています(Limら, 2007)。つまり、Zic3中胚葉エンハンサーはES細胞が内胚葉になるか、それ以外(中胚葉および外胚葉)になるかの選択に関わる可能性があります。
また、Zic3中胚葉エンハンサーのうち、どの部分が活性化しているか(DNAが露出して細胞の運命決定に関わるタンパク質が結合するか)もデータベースを利用して、検討しました(上の図の「DNA-seq」)。上の図からわかるように、ES細胞や中胚葉(mesoderm)と、胎仔の脳組織(fetal brain, forebrain) の間では違う部分が活性化しています。私たちがマウスで行ったレポーター実験では、一部の胚で脳の表面に近い部分でZic3中胚葉エンハンサーが活性化していました(下図)。これらのことから、「Zic3中胚葉エンハンサー」には外胚葉から生じる神経組織でも働く潜在的な活性があるのではないかと考えられました。
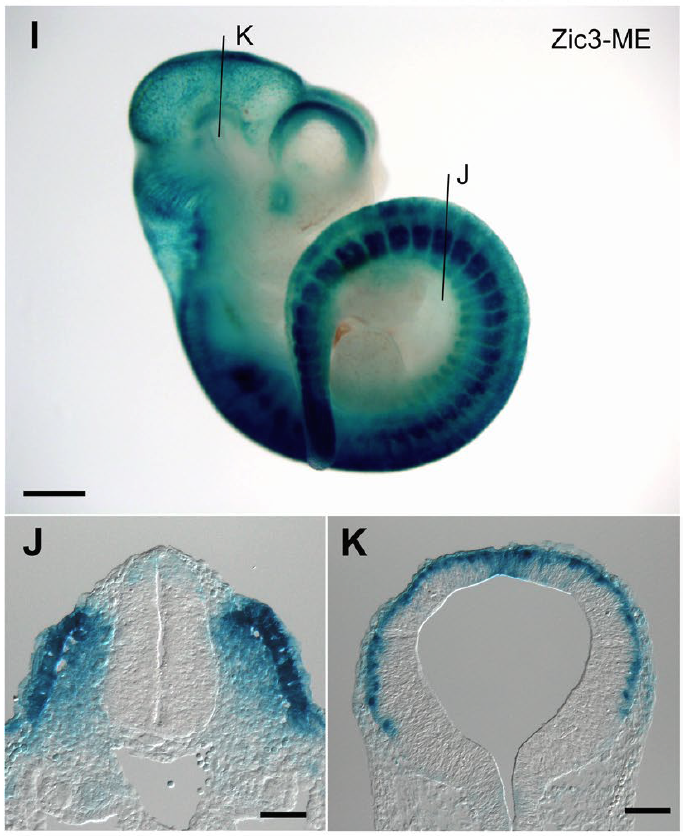
Zic3中胚葉エンハンサーが働いている。
体節(J)と脳の表面に近いところ(K)に
シグナルが見られる。
細胞の核にあるDNAの中には、どの遺伝子の情報を読み出し、どの遺伝子の情報を眠らせておくか、を決める領域(遺伝子発現調節領域)が数多く島のように散りばめられています。そして、それらの領域のDNAがメチル化されたり、近傍のヒストンがアセチル化やメチル化されたりするなど様々な修飾によって調節されていることがわかっていましたが、近年、上述のような1次元的な制御だけでなく、離れた場所の制御領域が立体的に近づくことでより複雑な遺伝子のオンオフの調節がなされるということが分かってきました。結合するタンパク質の性質などから、「Zic3中胚葉エンハンサー」もZic3遺伝子に近づくことで遺伝子発現を調節しているものと考えられます(下図)。
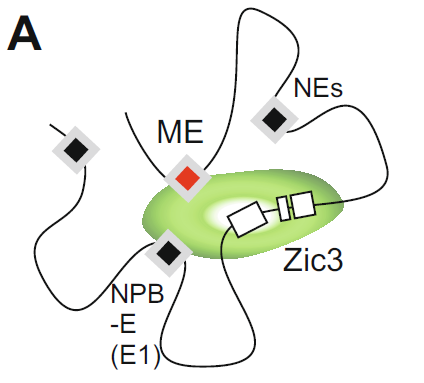
離れたところ(ヒトでは170kb)に存在している。
他のエンハンサー(NE、NPB-E)やZic3近傍の発現制御
領域と共にZic3の発現制御を行っている。
今回の研究で私たちが発見した「Zic3中胚葉エンハンサー」は受精卵から中胚葉がつくられ、さらに様々な細胞へと分化していく過程に必要な制御領域であることがわかりました。この領域を中心として様々な初期胚の細胞運命の決定に関わるシグナルやさまざまな転写因子が集合しZic3の遺伝子発現を制御している姿が見えてきました。これらの知見が幹細胞のふるまいの理解やその応用に役立つことを期待しています。
