
不安や抑うつの原因となる遺伝子が出生直後の脳のノルアドレナリン作動性神経の発達を制御していることを発見
Hatayama et al., Commun Biol (2022)
膜貫通タンパク質SLITRK1はこれまでの研究でトゥレット障害(チックが頻繁に現れる)や強迫性障害(特定の行動を繰り返してしまう)と関わる可能性が示されてきました。一方、大人のSLITRK1を持たないマウス(SLITRK1欠損マウス)では不安やうつ病のような症状が強くなることを私たちの以前の研究で報告しました(トゥレット症候群関連遺伝子の機能解明 (Katayama et al., 2009)) 。今回、私たちは「SLITRK1がマウスの脳の中で無くなることが、どのようにして不安・うつ様の症状を引き起こすのか」を知るために、脳の発達過程に注目して研究しました。
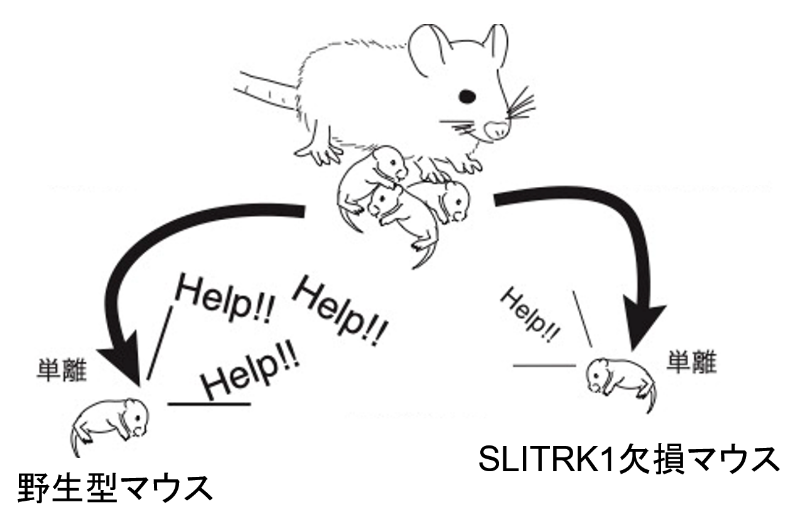
まず、SLITRK1欠損マウスの発達過程で、行動の異常がいつから観察されるのかを詳細に調べてみました。すると、最も早い症状は生後7日目に「母親マウスから離されたときに、助けを求める鳴き声が弱く少なくなる」(下図)ことであることがわかりました。マウスは生後19日目で離乳するので、7日目は新生児期に当たります。一方、大人のSLITRK1欠損マウスで観察される不安・うつ様の症状は生後5週から観察されました(マウスは約7週で性成熟します)。これらのことから、私たちはSLITRK1が遅くとも生後7日目から働き始め、脳の発達過程での働きが不安やうつ様の症状を引き起こすことに関わっているのであろうと推測しました。
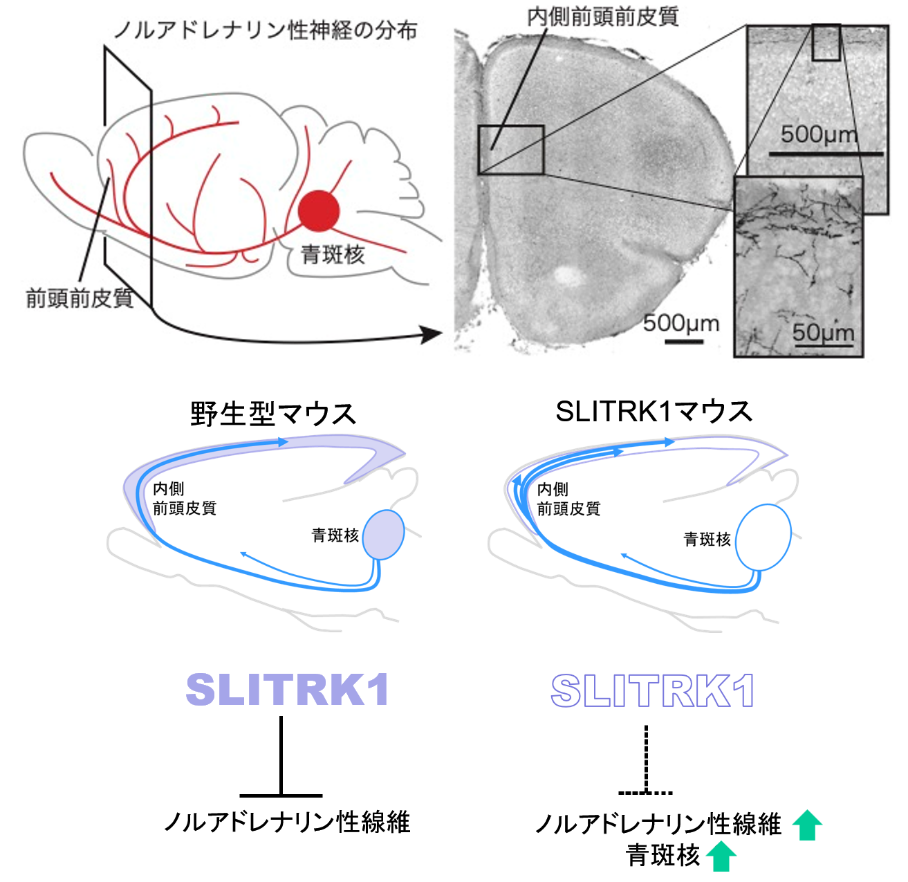
一方、ヒトの不安障害やうつ病の治療に使われる薬の多くはモノアミン(セロトニンやノルアドレナリンなど)を介した神経細胞間での情報のやりとりを調節します。そこで、私たちはSLITRK1欠損マウスで、モノアミンを放出する神経細胞の形や大きさ、脳の各部位でのモノアミンの含量を調べてみました。その結果わかったのは、生後3日目から7日目にかけて内側前頭前皮質でノルアドレナリンを放出する神経細胞(ノルアドレナリン性神経細胞)の神経突起が過剰に分布しており、ノルアドレナリン性神経細胞の細胞体がある青斑核のサイズが大きくなっていたことです(下図)。この時期の内側前頭前皮質ではノルアドレナリンの量も増えていました。同時にセロトニンが蓄えられている部分(セロトニン性神経細胞の神経突起にある膨らみ)が大きくなっていました。これには、この時期のノルアドレナリンはセロトニンが放出される部分にも作用して、その放出を抑えるはたらきがあることが関係しているものと考えられました。
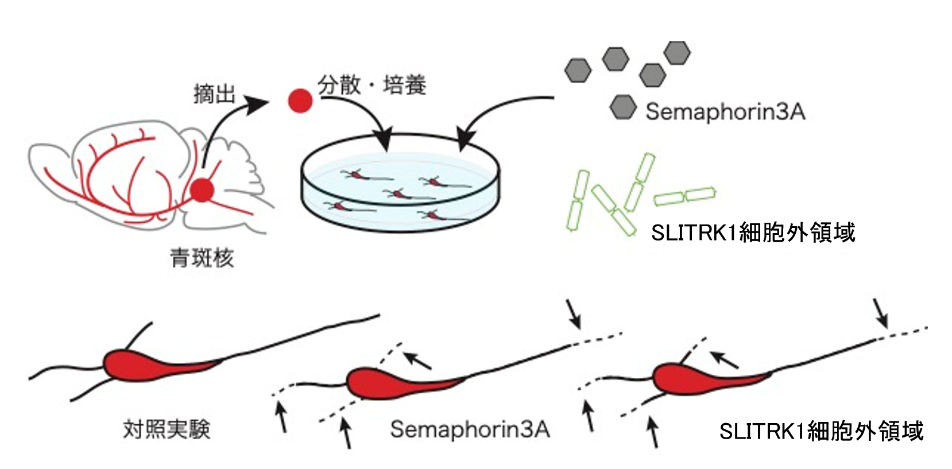
次にどのようにして、ノルアドレナリン性神経細胞の過剰な発達が起きるのかを調べることにしました。SLITRK1は細胞膜を貫通するタンパク質で細胞外に多く露出しています。生後7日目の脳では大脳皮質でも、ノルアドレナリン性神経細胞のある青斑核でもSLITRK1は作られています。青斑核の細胞だけを取り出して、培養液の入った培養皿の中においておくと神経突起を伸ばす様子が観察できます。この実験での無い青斑核のノルアドレナリン性神経細胞中のSLITRK1が無くなると、神経突起の形が変化することがわかりました。さらにSLITRK1タンパク質の細胞の外に出ている部分(SLITRK1細胞外領域)を作って、培養されている青斑核の神経細胞に作用させた場合には神経突起が伸びにくくなることがわかりました(下図)。
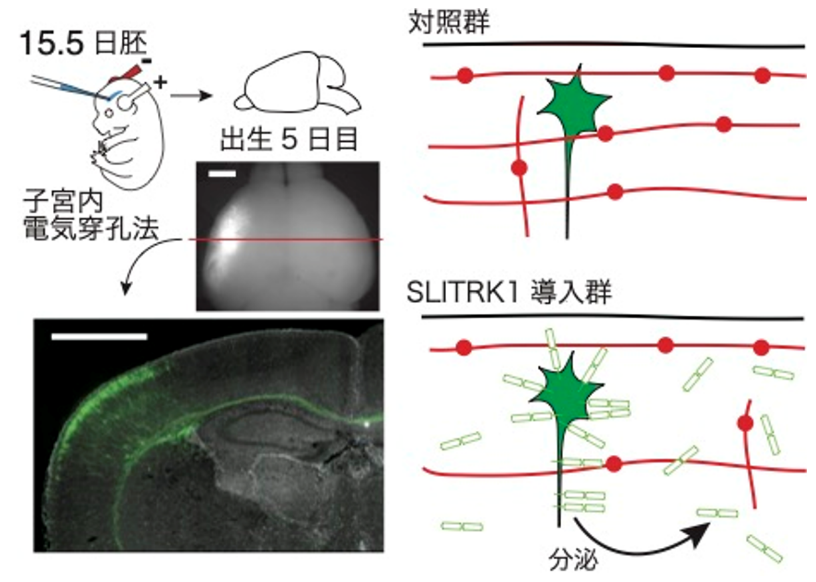
脳の中でもこのような活性があるのかを調べるために、胎仔期にたくさんのSLITRK1遺伝子を脳の中に注射して、大脳皮質に過剰にSLITRK1タンパク質を作り出してやると、ノルアドレナリン性神経細胞の突起が減ることもわかりました(下図)。これらのことから、SLITRK1タンパク質は細胞の中からも細胞の外からもノルアドレナリン性神経細胞の神経突起の発達を調節できることと、細胞の外から働いたときには突起が伸びるのを抑えることがわかりました。
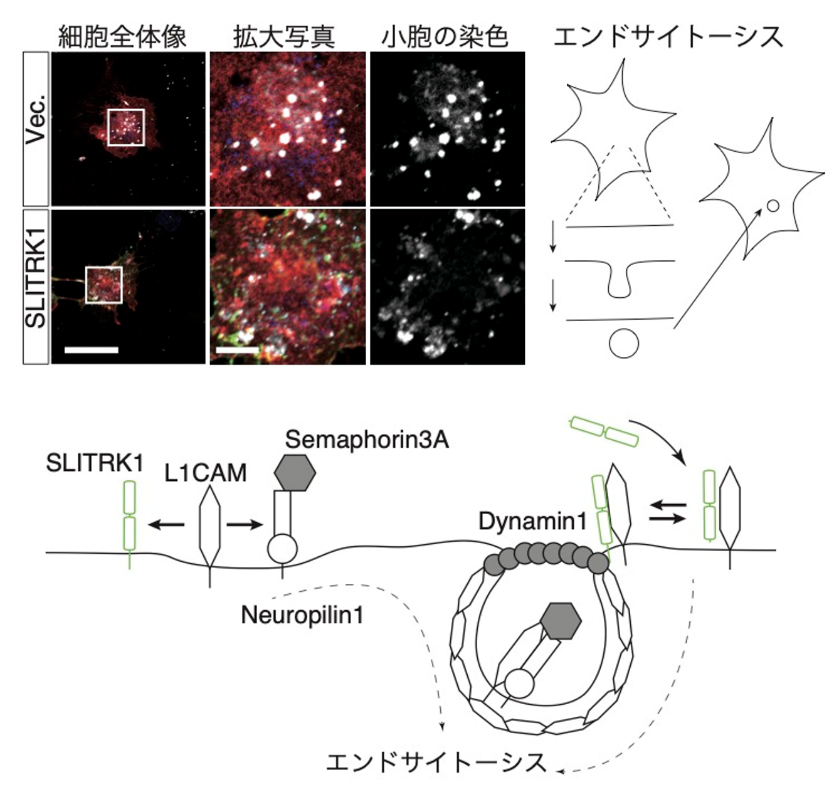
更に詳細にSLITRK1がどのようにして神経突起が伸びるのを抑えるのかを知るために、SLITRK1がどのタンパク質と結合するのかを調べました。出生直前のマウス脳を取り出して、SLITRK1に対する抗体を使ってSLITRK1を抽出したときに一緒に抽出されたタンパク質を調べて、それらとSLITRK1が結合するかどうかを調べたところ、Neurofascin(膜貫通タンパク質)やL1CAM(膜貫通タンパク質)、Dynamin(細胞内タンパク質)がSLITRK1と結合することがわかりました。これらのタンパク質のうち、L1CAMは神経突起の伸展を抑えるSemaphorinという分子と一緒に働くことが知られています。また、Dynaminは細胞膜がくびれて袋状になり、中に取り込まれる際(エンドサイトーシス)に働くことが分かっています。この結果を受けて、いくつかの実験を行った結果、SLITRK1は細胞突起の伸展を調節するSemaphorinのシグナルと関わることと、エンドサイトーシスの過程を調節することで神経突起の伸展を制御するものと考えられました(下図)。
今回の研究から、SLITRK1がノルアドレナリン性神経細胞の発達過程で重要な役割を持つことが明らかになりました。脳の中のノルアドレナリン性神経細胞は外界に対して注意が必要な状況で盛んに活動して脳全体の調子を整え、ほかのモノアミン性神経系とともに「覚醒」の状態をつくりだすことに働いているものと考えられています。この研究の結果、新生仔期のノルアドレナリンシグナルの過剰はその後の神経回路の性質をどのように変えるのか(神経回路の可塑性)という新たな課題が生まれました。この問題に答えることが脳の発達のメカニズムを理解する上で重要であろうと考えています。
